Questions fréquentes concernant le virus de la PPA détecté à Barcelone
25-Mai-2026 (il y a 1 mois 16 jours)
Comment classe-t-on les virus de la PPA ?
Le virus de la peste porcine africaine (VPPA) a été classé en différents génotypes, en fonction de la séquence du gène B646L, qui code pour la protéine p72, composant principal du virus. Sur la base de cette séquence, jusqu’à 24 génotypes distincts ont été décrits, circulant sur le continent africain.
En dehors de l'Afrique, seule la circulation des génotypes suivants a été signalée :
- Génotype I : circulant actuellement en Asie, et responsable de la pandémie qui a débuté dans les années 60.
- Génotype II : le plus répandu dans la pandémie actuelle, qui a débuté en Géorgie en 2007.
Le laboratoire de référence européen a proposé une classification en groupes génétiques au sein du génotype II sur la base des différences de séquences dans six régions variables du génome du virus.
| CVR | IGR I73R/I329L | O174L |
| K145R | IGR MGF505 9R/10R | IGR I329L-I215L |
Sur la base des différences observées dans ces régions, 28 groupes génétiques au total avaient été décrits, le groupe génétique n° 1 étant représenté par la souche Georgia2007.
La nomenclature proposée pour la souche découverte à Barcelone est SP25WB2611, abrégée en Sp25.
La souche découverte à Barcelone (Sp25) est différente de celles connues jusqu'à présent et a donc été classée dans un nouveau groupe génétique, le n° 29.
Certains groupes génétiques n’ont été isolés que chez le porc domestique, d’autres uniquement chez le sanglier et d’autres encore chez les deux espèces. Il existe des groupes génétiques, comme le n° 3 ou le n° 19, qui sont présents dans différents pays, mais la plupart n’ont été détectés que dans une zone géographique spécifique. De même, il existe des régions où circule plus d'un groupe génétique, comme par exemple en Fédération de Russie (n° 1, n° 2, n° 3, n° 4 et n° 7) ou en Roumanie (n° 3, n° 7, n° 19, n° 21, n° 22 et n° 24).
Le génome de la souche Sp25 présente une grande délétion, d'environ 10 Kb, à l'extrémité gauche du génome, ainsi qu'un total de 27 SNP (mutations ponctuelles produisant des changements d'un nucléotide dans la séquence du virus, (de l'anglais Single Nucleotide Polymorphisms) par rapport à la souche Georgia 2007.
Comment les séquences disponibles dans la banque de gènes Genebank, auxquelles les nouvelles souches sont comparées, sont-elles réparties dans le temps et dans l'espace ?
Bien que de nombreuses souches de la PPA aient été isolées en Europe depuis 2007, le nombre de séquences obtenues reste relativement faible.
La connaissance de la répartition géographique des groupes génétiques dépend du nombre d'échantillons séquencés et publiés dans chaque pays. Dans de nombreux pays touchés par la PPA, les séquences ne sont pas séquencées et/ou les résultats ne sont pas publiés.
Les analyses de la séquence de la souche Sp25 réalisées à l'IRB et au BSC, à Barcelone, montrent sa proximité phylogénétique avec des souches isolées lors de foyers survenus dans des pays européens après l'introduction de la souche géorgienne de 2007 sur le continent (Figure 1). La souche Sp25 appartient à une lignée qui diverge très tôt et est donc très différente de celles à l'origine des épidémies les plus récentes en Europe centrale et occidentale. Comme le montre l'arbre, cette séquence présente une plus grande proximité avec les virus détectés en Russie en 2019 qu'avec les souches isolées en Europe centrale et occidentale, mais elle reste néanmoins très différente de toute souche dont le génome complet a été séquencé, y compris celles isolées en Russie, dont elle diffère par la délétion mentionnée et par plus de 20 SNP.

On dit que le virus de la PPA est un virus à ADN très gros et très stable. Combien de changements un isolat de PPA doit-il présenter pour être considéré comme une souche distincte ?
Dans le cadre du VPPA en Europe, le laboratoire de référence européen considère qu’un isolat viral constitue une souche distincte des autres lorsqu’il est classé dans un nouveau groupe génétique.
Le nombre de SNP détectés nous permet d'évaluer la distance temporelle d'une souche par rapport à la souche « originale » Georgia2007. Le VPPA est un virus à ADN double brin et, par conséquent, un virus très stable. Compte tenu du faible taux de mutation du virus, les différents SNP présentés par Sp25 doivent s'être accumulés sur une longue période depuis sa divergence par rapport à Georgia2007. Selon les dernières estimations, cette période pourrait correspondre à environ 10 à 15 ans de divergence évolutive (estimation réalisée par le Dr Toni Gabaldón, IRB).
Si ce virus est si stable, la délétion de 10 000 paires de bases observée dans la souche responsable de l'épidémie de Barcelone peut-elle se produire naturellement ?
La dynamique évolutive naturelle du virus génère ce type de délétions génétiques. Au sein du génotype II, des délétions dans la même région, avec une étendue et une atteinte des gènes différentes, avaient déjà été décrites tant en Afrique (Ambagala et al., 2023) qu'en Europe (Zani et al., 2018 ; Torresi et al., 2025).
Cette délétion est utilisée dans certains candidats vaccins. Cela signifie-t-il qu'elle ne peut pas se produire naturellement ?
Ce type de délétions dans le génome du VPPA se produit naturellement et peut également être généré en laboratoire pour créer certains candidats vaccins. Il convient de souligner qu'aujourd'hui, lors du protocole de développement d'un candidat vaccin en laboratoire, on inclut dans la plupart des cas un marqueur exogène dans le génome du virus (pour des raisons techniques) qui permet de le différencier de la souche originale. Par conséquent, les candidats vaccins sont facilement identifiables par séquençage. La souche Sp25 ne présente aucun marqueur exogène. De plus, la plupart des candidats vaccins sont générés à partir d'un petit groupe de souches de laboratoire très similaires entre elles et à la première souche isolée lors de l'épidémie actuelle, Georgia2007.
La recombinaison entre différentes souches de PPA est-elle fréquente ?
Pour que ces recombinaisons aient lieu, une cellule de l'hôte doit être infectée simultanément par deux souches différentes. Ce phénomène n'est pas aussi courant que chez les virus à ARN à génome fragmenté (comme c'est le cas du virus de la grippe), mais il se produit bel et bien avec le VPPA. En Chine, des recombinaisons entre les génotypes I et II ont été signalées (Zhao et al., 2023) et ce même virus recombinant s'est propagé vers d'autres pays (Russie et Vietnam). Logiquement, plus il y a de souches différentes en circulation dans une même zone, plus la probabilité que de telles recombinaisons se produisent est élevée. Le profil des mutations détectées dans la souche de Collserola est réparti sur l'ensemble du génome, et non dans une région limitée, ce qui rend improbable une origine par recombinaison.
On entend beaucoup dire que la souche détectée à Barcelone n'est pas très virulente. Est-ce vraiment le cas ?
La délétion identifiée dans la souche Sp25 pourrait être associée à une virulence moindre, comme cela a été démontré pour d'autres souches présentant des délétions affectant cette région. Cependant, il existe des exemples de souches très virulentes présentant des délétions similaires, ce qui rend cette interprétation non concluante. La virulence de la souche Sp25 ne pourra être évaluée qu'après la réalisation d'une infection expérimentale chez des porcs ou des sangliers, en conditions contrôlées.
La seule certitude dont nous disposons à l'heure actuelle est qu'il s'agit d'une souche mortelle chez les sangliers et que le profil de la maladie observé est comparable à celui décrit lors d'autres épidémies européennes causées par des souches classées comme hautement virulentes.
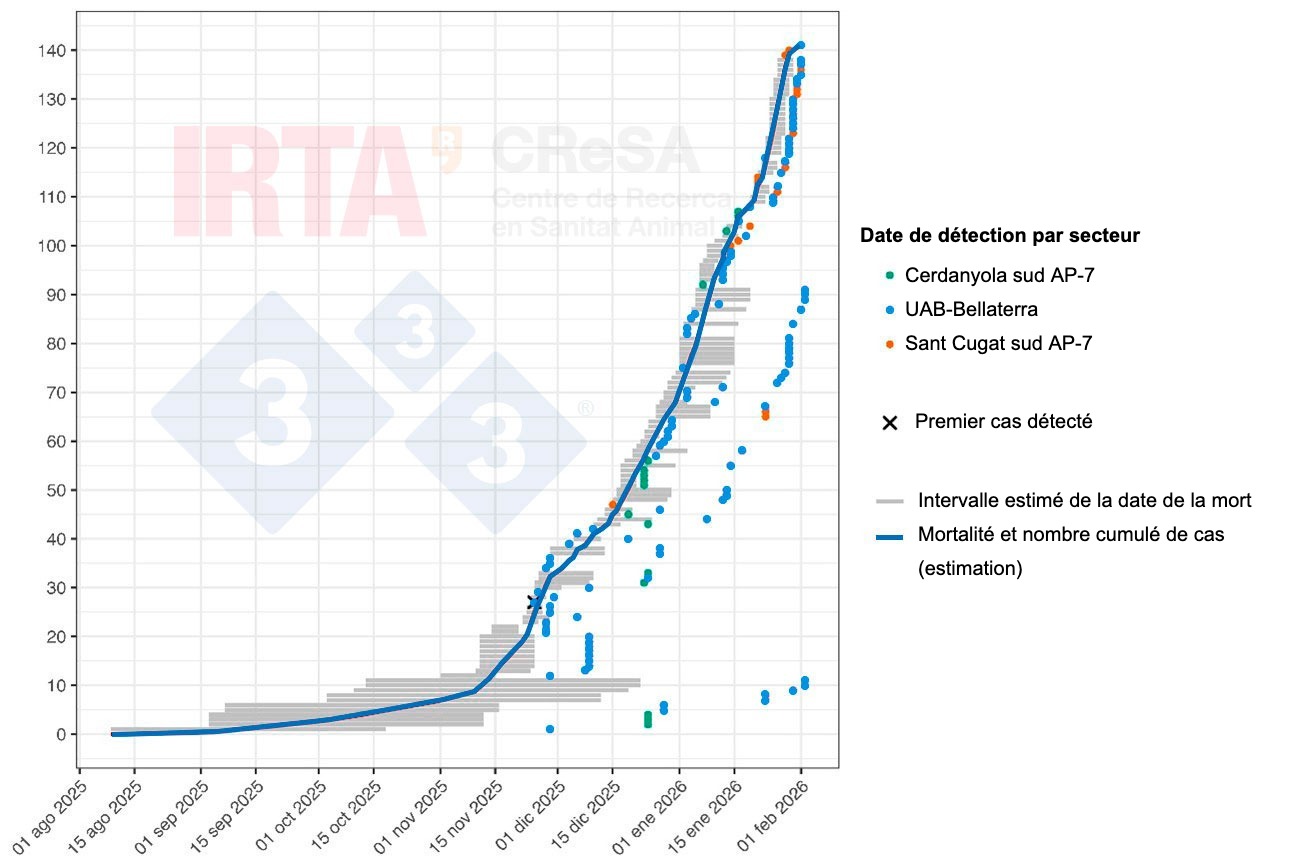
La figure 2 présente une reconstitution chronologique des cas chez les sangliers dans le cadre de la présente épidémie de PPA, en tenant compte du fait que la date de découverte du cadavre ne coïncide généralement pas avec la date de la mort de l'animal. Pour chaque cas, un intervalle de temps plausible pour la mort du sanglier est présenté, estimé à partir de l'état de conservation du cadavre. Ainsi, bien que la date exacte des premières morts ne puisse être déterminée avec précision, les intervalles estimés pour les premiers cadavres indiquent que la mortalité associée à l'épidémie a commencé plusieurs mois avant la détection du premier cas, probablement vers le mois de septembre. Le VPPA a donc circulé sans être détecté pendant une période initiale, probablement à un niveau faible mais soutenu, jusqu'à ce qu'il soit détecté. La phase initiale de l'épidémie, à progression lente, correspond à l'introduction et à l'établissement du VPPA dans la population de sangliers, suivie d'une phase épidémique au cours de laquelle on observe un changement de pente de la courbe. Il convient de souligner que cette évolution de l'épidémie n'est pas très éloignée de celle observée lors d'épidémies récentes en Europe.